Последовательные ступени развития лошадиного ряда
Фенакод даёт нам ясное представление о древнейшем типе строения непарнокопытных четвероногих, однако сам он не стоит в ряду прямых предков лошадиного ряда, так как одновременно с ним на Земле уже существовали формы с утраченными боковыми пальцами — прямые предшественники позднейших однокопытных.
Наиболее древним членом этого ряда можно считать раннетретнчного эогиппуса. Это было небольшое животное ростом с лисицу, имевшее по 4 пальца на передних конечностях и по 3 на задних; при ходьбе его боковые пальцы должны были касаться земли. После эогиипуса идёт несколько форм с трёхпалыми конечностями (IV палец уже утрачен). Далее появляется более крупная трёхпалая форма — моногиппус, у которой средний палец развит значительно сильнее, а боковые уже не соприкасаются с гладкой поверхностью земли.
Следующие члены ряда — животные ещё более крупных размеров, у которых боковые пальцы становятся явно бесполезными рудиментами, хотя и остаются заметными снаружи. Наконец, верхнетретичный плиогиппус оказывается уже однокопытным животным, относительно близким к современным видам семейства лошадей — наиболее крупным представителям этого ряда.
Параллельно с недоразвитием боковых пальцев и увеличением размеров тела у членов лошадиного ряда шло и изменение зубного аппарата. У древнейших форм коренные зубы были бугорчатые, у современных лошадей они имеют плоскую жевательную поверхность и складчатое строение, а промежуточные члены ряда дают различные переходы между этими двумя крайними типами.
Чем же было обусловлено направление процесса, который с такой неуклонной последовательностью выразился в постепенных переходах от мелкого многопалого эогиппуса к современным длинноногим однокопытным лошадям, от многобугорчатых коренных зубов к зубам цилиндрической формы, способным перетирать сухое сено и твёрдые зерновые корма?
Этот вопрос получил блестящее разрешение ещё в 70-х годах прошлого столетия в работах гениального русского палеонтолога Владимира Онуфриевича Ковалевского.
Более древние животные в ряду предков лошадиного ряда не были степными. И по их строению, и по характеру отложений, в которых были найдены их остатки, видно, что они жили в условиях тёплого и влажного климата и питались сочным растительным кормом (вспомним подобные же бугорчатые зубы наших свиней и их корм).
При небольших размерах среди густой растительности этим животным не требовалось быстроты и неутомимости бега, которые необходимы обитателям открытых пространств — современным диким однокопытным, лишённым возможности укрыться в норе и в густых зарослях. В этих условиях сравнительно короткие трёхпалые или четырёхпалые ноги вполне удовлетворяли потребности животного, тем более что в случае надобности оно могло ускорить свой бег, передвигаясь скачками при помощи сгибания и разгибания своего туловища (вспомним движения кошки).
Геологические перемены, которые наступили в середине третичного периода и сопровождались поднятием высоких горных хребтов, вызвали изменение климатических условий на больших пространствах суши, а вместе с тем и изменение растительного покрова. В странах, которые оказались отделёнными от морей высокими горными хребтами, климат стал более континентальным и лесная растительность сменилась травянистыми степями.
В этих условиях для крупных копытных животных был уже отрезан путь к выработке способности рыть норы и укрываться в них от опасности, как это делают грызуны и мелкие хищники, и единственным средством спасения остался для них быстрый бег. Но при крупных размерах тела уже утрачивается гибкость позвоночника, позволяющая мелким четвероногим двигаться скачками, и быстрота передвижения начинает зависеть только от ног.
В этих новых условиях жизненное преимущество получили животные с более длинными ногами и с возможно более сокращённой поверхностью опоры, то есть уменьшенным количеством пальцев (ведь недаром олицетворением медлительности для нас служит брюхоногий моллюск — улитка, и недаром мы сами, опираясь при ходьбе на всю ступню, при беге обязательно поднимаемся на кончики пальцев).
С переходом к жизни в степях изменился и характер питания копытных: животные растительноядные с бугорчатыми зубами под неумолимым действием естественного отбора постепенно переродились в животных травоядных, обладающих складчатыми зубами и способных поедать растительность и после того, как она будет выжжена солнцем и засохнет на корню.
Таким образом, на основании подлинных палеонтологических документов оказалось возможным не только установить филогенетический ряд семейства лошадей, но и выяснить причины, почему развитие этой группы пошло в таком именно направлении.
Классические работы В. О. Ковалевского по изучению вымерших групп копытных положили начало экологическому направлению в палеонтологии, которое видит в ископаемых остатках не только мёртвые кости, но и части действовавших в своё время живых организмов, строение которых соответствовало условиям их существования и их образу жизни.
Если вам доведётся видеть полный скелет лошади, вы можете заметить, что приспособление конечностей к длительному и быстрой бегу не ограничивается утратой боковых пальцев, а выражается ещё в упрощении строения предплечья (рис. 463) и голени: локтевая кость срослась с лучевой, а малая берцовая обратилась в небольшой придаток большой берцовой кости.
В результате всех этих изменений и при отсутствии ключиц для лошади недоступны вращательные движения в плечевом суставе и повороты её однопалой кисти (заметим, что дрессированные цирковые лошади, поднявшись на задние ноги и кланяясь на аплодисменты зрителей, не могут сделать приветственных движений передними конечностями и только перебирают ими в вертикальной плоскости).
Но как раз именно такая скованность движений придаёт высоким ногам лошади устойчивость и крепость, необходимые при быстром беге по твёрдому грунту открытых степей, в одомашненном состоянии закрепила за лошадьми их первостепенное значение в качестве верховых и упряжных животных.
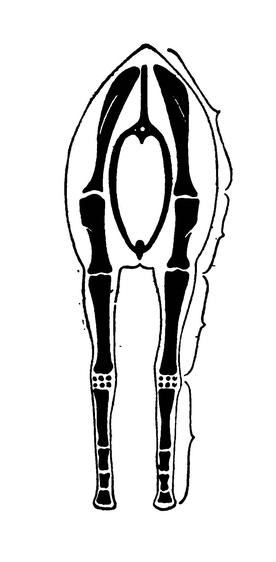
Рис. 463. Расположение звеньев передних конечностей лошади
Придающее им достаточную устойчивость и вместе с тем ограничивающее их подвижность маятникообразными движениями в одной плоскости при ходьбе и беге (вид спереди; схема).
← Предыдущая статья
ФенакодСледующая статья →
Оскудение фауны копытных
Образы животных в пословицах и поговорках
Волка на собак в помощь не зови.
Не велик сверчок, а громко поёт.
Рыба с головы гниёт.
Оружие животных

Как нож в воде
Животные, чрезвычайно приспособленные по окраске и форме к окружающей среде, ...
Рак-щелкун
Относящийся к креветкам, с разной величиной клешнёй рак-щелкун водится в ...Книги

Язык лошадей. Образ жизни, поведение, формы общения
Душевное и физическое здоровье лошади, как минимум, наполовину зависит от ...